Spierspoeltjes
Spierspoeltjes zijn vooral te vinden tussen de spiervezels van spieren die verantwoordelijk zijn voor de lichaamshouding. Het zijn zintuigjes van ongeveer 7 millimeter lang die gewikkeld zijn rond dwarsgestreepte spiervezels.





Deze mechanoreceptoren zenden informatie door naar de hersenen over de rekbaarheidstoestand van de spieren. Ze wisselen daar in een constante stroom informatie over uit. Een spierspoeltje bestaat uit intrafusale vezels. Een spierspoeltje heeft twee sensorische (afferente) en een motorische zenuwvezel (efferente). Informatie van sensorische zenuwcellen geven via schakelcellen informatie door aan motorische zenuwcellen, de reflexboog. De buitenste weefsellaag van het spierspoeltje bestaat uit afgeplatte, bindweefselcellen. In de ruimte rondom de zenuwvezels zit een met vloeistof gevulde holte.
Spierspoeltjes hebben een uitrekbaar middenstuk en samentrekbare uiteinden. Hiermee kunnen ze het samentrekken en uitrekken van een spier regelen voordat er door het lichaam actie wordt ondernomen. Dit kan door de nauwe samenwerking met de hersenen. Als spieren een bepaalde houding moeten volhouden werken de spoeltjes als fixatiezintuig.
Rol van het spierspoeltje tijdens gecontroleerde bewegingen
bewerkenOm het belang van het gamma-motoneuronen te benadrukken, dient gerealiseerd te worden dat 31% van alle motoneuronen naar de spier type A-gamma-motoneuronen zijn. Als signalen vanuit de cortex (of ander hersengebied) naar de alfa-motoneuronen worden gezonden, worden tevens de gamma-motoneuronen gestimuleerd. Deze co-activatie heeft intrafusale spiercontractie tot gevolg. De gelijktijdige contractie van intra- (in het spierspoeltje) en extrafusale (buiten het spierspoeltje) spiervezels heeft twee belangrijke doelen:
- de lengte van de receptor van de spierspoel wordt gelijk gehouden en daardoor blijft de receptor altijd exciteerbaar (prikkelbaar);
- de spierspoel kan zo te allen tijde spiercontracties bijstellen.[1]
Bouw
bewerkenSpierspoelen bestaan uit 5-14 spiervezels, waarvan er drie soorten zijn: dynamische nucleaire zakvezels (zak1-vezels), statische nucleaire zakvezels (zak2-vezels) en nucleaire kettingvezels. De nucleï zitten bij de zakvezels geconcentreerd in en soort zak (uitstulping) in het midden van de vezel. Bij de kettingvezels zitten ze verspreid over de gehele lengte van de vezel. De kettingvezels zijn dunner en korter dan de zakvezels.
Primaire sensorische vezels van type Ia (grote diameter) zitten spiraalvormig rond alle intrafusale (in het spierspoeltje) spiervezels en eindigen nabij het midden van elke vezel met een "annulospiraal" uiteinde. Secundaire type II sensorische vezels (gemiddelde diameter) eindigen grenzend aan de centrale gebieden van de statische zak- en kettingvezels met "flower spray"-uiteinden die sproeivormig of ringvormig kunnen zijn en zich in smalle banden aan beide zijden van de ketting- of zakvezel uitspreiden. Deze vezels verzenden informatie via rekgevoelige mechanosensitieve ionkanalen van de axonen (zenuwvezels). De type Ia sensorische vezels hebben een snelle geleiding van 70 tot 120 m.s-1. De langzaam geleidende type II sensorische vezels hebben een geleidingssnelheid van 36 tot 72 m.s-1.
Het motorische deel van het spoeltje heeft motoneuronen (motorneuronen): tot een dozijn gamma-motoneuronen, ook wel fusi-motoneuronen genoemd. Deze activeren de intrafusale spiervezels in het spoeltje. Gamma-motoneuronen activeren alleen spiervezels binnen het spoeltje, terwijl bèta-motoneuronen spiervezels zowel binnen als buiten het spoeltje activeren. Activering van de neuronen veroorzaakt een samentrekking en verstijving van de beide uiteinden van de spierspoeltje spiervezels.
Gamma-motoneuronen worden geclassificeerd als statisch of dynamisch afhankelijk van het type spiervezels die ze innerveren en hun effecten op de reacties van de Ia- en II-sensorische neuronen die het centrale, niet-samentrekbare centrale (equatoriale) deel van de spierspoel innerveren.
- De statische axonen innerveren de ketting- of statische zak2-vezels. Ze verhogen de prikkelsnelheid van Ia- en II-sensorische zenuwvezels (afferenten) bij een bepaalde spierlengte.
- De dynamische axonen innerveren de intrafusale (in het spierspoeltje) spiervezels van de zak1. Ze verhogen de rekgevoeligheid van de Ia-sensorische zenuwvezels (afferenten) door de intrafusale vezels van zak1 te verstijven.
-
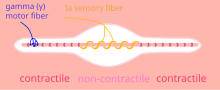 Spierspoeltje (fiber = vezel; contractile = samentrekbaar)
Spierspoeltje (fiber = vezel; contractile = samentrekbaar) -
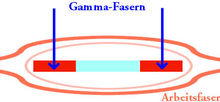 Gamma-vezel. Signalen komen van de extrapiramidale routes in de hersenstam via gamma-motoneuronen
Gamma-vezel. Signalen komen van de extrapiramidale routes in de hersenstam via gamma-motoneuronen -
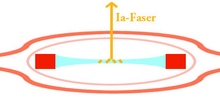 1a-vezel. De uiteinden van de spierspoelvezels trekken zich dan samen, de uitrekking van het midden van het spierspoeltje wordt geregistreerd en doorgegeven aan het ruggenmerg
1a-vezel. De uiteinden van de spierspoelvezels trekken zich dan samen, de uitrekking van het midden van het spierspoeltje wordt geregistreerd en doorgegeven aan het ruggenmerg -
 Alfa-vezel. De skeletspiervezels trekken samen, waardoor het spierspoeltje korter wordt
Alfa-vezel. De skeletspiervezels trekken samen, waardoor het spierspoeltje korter wordt




Functie
bewerken
Eenvoudige reflexen
bewerkenReflexen kunnen vrij eenvoudig verlopen, zoals monosynaptische reflexen (waarbij slechts één motorisch neuron betrokken is) van de skeletspieren. Men spreekt ook wel van spinale reflexen. Hierbij is sprake van slechts één schakel of synaps in het ruggenmerg. Een belangrijk rol spelen hier de proprioreceptoren. Deze liggen in de gewrichten, pezen en spierspoeltje van een spier. Spinale reflexen van de spieren verlopen zeer snel en worden ook wel spier-eigenreflexen genoemd. De receptor ligt hier namelijk in het reagerende orgaan zelf: de spier reageert als gevolg van de rekking van de spier zelf. Veel van deze reflexen ontstaan via goed beschreven neuronale circuits. Vaak veroorzaakt uitrekking van die spier dan een samentrekking.
Een bekende reflex is de kniepeesreflex. Deze reflex ontstaat door een plotselinge rek van de bovenste dijbeenspier bijv. door het uit balans raken van het lichaam, maar ook tijdens lopen, springen e.d. Peesreflex is dan ook een onjuiste naam; spierrekkingsreflexen dekt de lading beter. De actieve samentrekking van de bovenste dijbeenspier is dus een reactie op de eerdere passieve rekking ervan. Tegelijkertijd gaat dit gepaard met een ontspanning van de antagonistspier. Het laatste gebeurt via een remmend tussenneuron in het ruggenmerg. Dit voorbeeld illustreert dat zelfs bij eenvoudige reflexen sprake kan zijn van meerdere zenuwcellen of interneuronen tussen het sensorische en motorische neuron. Deze dienen om een gecoördineerde beweging waarbij meerdere spieren zijn betrokken tot stand te brengen.
Complexe reflexen
bewerkenMeer complexe reflexen verlopen via meerdere synapsen. Zo lopen de reflexen van het autonome zenuwstelsel via een extra synaps in het ganglion, een streng zenuwcellen die buiten het ruggenmerg loopt. Dit is dus een extra station dat gepasseerd wordt op weg naar het doelorgaan, bijvoorbeeld een speekselklier, gladde spier of de hartspier. De prikkels die deze reflexen veroorzaken zijn bijvoorbeeld pijnprikkels, of prikkeling van mechanoreceptoren en chemoreceptoren van inwendige organen zoals de long, maag-darmkanaal en bloedvaten. De eindorganen worden bovendien door twee soorten motorische zenuwvezels aangestuurd, een orthosympathisch en een parasympathisch deel. De reflexen die via meerdere synapsen lopen, zijn langzamer en worden ook wel vreemde reflexen genoemd. Dit wil zeggen dat de receptor die de prikkel ontvangt hier ruimtelijk gescheiden is van het reagerende orgaan. Een voorbeeld is de buigreflex waarbij prikkeling van de voetzool leidt tot buiging van gewrichten in het been. Andere voorbeelden zijn de corneareflex, hoesten en niezen.
Rekreflex
bewerkenWanneer een spier wordt uitgerekt, reageren de primaire type Ia-sensorische vezels van het spierspoeltje op zowel veranderingen in spierlengte als snelheid en brengen deze activiteit over naar het ruggenmerg in de vorm van veranderingen in de snelheid van actiepotentialen. Op dezelfde manier reageren de secundaire type II sensorische vezels op veranderingen in spierlengte (maar met een kleinere snelheidsgevoelige component) en zenden dit signaal naar het ruggenmerg. De Ia-sensorische signalen worden monosynaptisch verzonden naar vele alfa-motoneuronen van de receptordragende spier. Hoe minder spiervezels door een α-motoneuron worden geïnnerveerd, hoe fijner de beweging kan worden afgestemd: in de oog- en vingerspieren innerveert een motoneuron ongeveer 100 spiervezels, in andere spieren tot 2000. De reflexmatig opgewekte activiteit in de alfa-motoneuronen wordt vervolgens via hun motorische zenuwcellen doorgegeven aan de extrafusale (buiten het spierspoeltje) vezels van de spier, die kracht genereren en daardoor weerstand bieden aan de rek. Het Ia-sensorische signaal wordt ook polysynaptisch overgedragen via interneuronen (Ia-remmende interneuronen), die alfa-motoneuronen van antagonistische spieren remmen, waardoor ze ontspannen.
Gevoeligheidswijziging
bewerkenDe functie van de gamma-motoneuronen is niet het aanvullen van de spiercontractiekracht die door de extrafusale vezels wordt geleverd, maar het wijzigen van de gevoeligheid van de sensorische vezels van het spierspoeltje voor rek. Bij het vrijkomen van acetylcholine door het actieve gamma-motoneuron trekken de uiteinden van de intrafusale (in het spierspoeltje) spiervezels samen, waardoor de niet-contractiele centrale gedeelten worden verlengd. Dit opent rekgevoelige ionkanalen in de sensorische uiteinden, wat leidt tot een instroom van natriumionen. Dit verhoogt het rustpotentiaal van de uiteinden, waardoor de kans op prikkelafgifte van het actiepotentiaal wordt vergroot, waardoor de rekgevoeligheid van de sensorische spierspoeltjesvezels toeneemt.
Hoe controleert het centrale zenuwstelsel gamma-motoneuronen? Het was moeilijk om tijdens normale beweging opnames te maken van gamma-motoneuronen, omdat ze zeer kleine zenuwvezels hebben. Er zijn verschillende theorieën voorgesteld, gebaseerd op opnames van sensorische spierspoeltjesvezels.
- 1) Alfa-gamma co-activatie. Hier wordt gesteld dat gamma-motoneuronen parallel met alfa-motoneuronen worden geactiveerd om de prikkelafgifte van sensorische spierspoeltjesvezels in stand te houden wanneer de extrafusale (buiten het spierspoeltje) spieren korter worden.[2]
- 2) Gamma-motoneuronset: Gamma-motoneuronen worden geactiveerd afhankelijk van de nieuwigheid of moeilijkheidsgraad van een taak. Terwijl statische gamma-motoneuronen continu actief zijn tijdens routinematige bewegingen zoals voortbeweging, worden dynamische gamma-motoneuronen doorgaans meer geactiveerd tijdens moeilijke taken, waardoor de sensorische Ia-strekgevoeligheid toeneemt.[3]
- 3) Gamma-motoneuronsjabloon van beoogde beweging. Statische gamma-activiteit is een "temporaalsjabloon" van de verwachte verkorting en verlenging van de receptordragende spier. Dynamische gamma-activiteit wordt abrupt in- en uitgeschakeld, waardoor de gamma-motoneuronen gevoelig worden voor het begin van spierverlenging en het afwijken van het beoogde bewegingstraject.[4]
- 4) Doelgerichte voorbereidende controle. Dynamische gamma-activiteit wordt tijdens de bewegingsvoorbereiding proactief aangepast om de uitvoering van de geplande actie te vergemakkelijken. Als de beoogde bewegingsrichting bijvoorbeeld verband houdt met rek van de spoeltjesdragende spier, wordt de sensorische Ia en rekreflexgevoeligheid van deze spier verminderd. Gamma-motoneuronencontrole maakt daarom de onafhankelijke voorbereidende afstemming van spierstijfheid mogelijk op basis van taakdoelen.[5]
Klinische betekenis
bewerkenNa beroerte of dwarslaesie bij mensen ontstaat vaak spastische hypertonie (spastische verlamming), waarbij de rekreflex in de buigspieren van de armen en de strekspieren van de benen te gevoelig is. Dit resulteert in abnormale houdingen, stijfheid en contracturen. Hypertonie kan het gevolg zijn van overgevoeligheid van alfa-motoneuronen en interneuronen voor de sensorische signalen Ia en II.[6]
- ↑ http://www.zowerkthetlichaam.nl/798/spierspoelen/
- ↑ Vallbo AB, al-Falahe NA (February 1990). Human muscle spindle response in a motor learning task. J. Physiol. 421: 553–68. PMID 2140862. PMC 1190101. DOI: 10.1113/jphysiol.1990.sp017961.
- ↑ Prochazka, A. (1996). Exercise: Regulation and Integration of Multiple Systems. American Physiological Society, New York, "Proprioceptive feedback and movement regulation", 89–127. ISBN 978-0195091748.
- ↑ Taylor A, Durbaba R, Ellaway PH, Rawlinson S (March 2006). Static and dynamic gamma-motor output to ankle flexor muscles during locomotion in the decerebrate cat. J. Physiol. 571 (Pt 3): 711–23. PMID 16423858. PMC 1805796. DOI: 10.1113/jphysiol.2005.101634.
- ↑ Papaioannou, S., Dimitriou, M. (2021). Goal-dependent tuning of muscle spindle receptors during movement preparation. Sci. Adv. 7 (9): eabe0401. PMID 33627426. PMC 7904268. DOI: 10.1126/sciadv.abe0401.
- ↑ Heckmann CJ, Gorassini MA, Bennett DJ (February 2005). Persistent inward currents in motoneuron dendrites: implications for motor output. Muscle Nerve 31 (2): 135–56. PMID 15736297. DOI: 10.1002/mus.20261.